Manganese (Mn) is an essential plant mineral nutrient, playing a key role in several physiological processes, particularly photosynthesis. Manganese deficiency is a widespread problem, most often occurring in sandy soils, organic soils with a pH above 6 and heavily weathered, tropical soils. It is typically worsened by cool and wet conditions (Alloway 2008). Numerous crop species have been reported to show high susceptibility to Mn deficiency in soils, or a very positive response to Mn fertilization, including cereal crops (wheat, barley and oats), legumes (common beans, peas and soybean), stone fruits (apples, cherries and peaches), palm crops, citrus, potatoes, sugar beets and canola, among others. The impact of Mn deficiencies on these crops includes reduced dry matter production and yield, weaker structural resistance against pathogens and a reduced tolerance to drought and heat stress.
Manganese has a relatively low phloem mobility in plants, and as a result, typical leaf symptoms of Mn deficiency first develop in younger leaves. The critical concentration for Mn deficiency is generally below 20 ppm dry weight in fully expanded, young leaves. In the case of dicots, Mn deficiency first results in pale mottled leaves, followed by typical interveinal chlorosis. Under severe Mn deficiency dicots may also develop a number of brownish spots. In cereals, Mn deficiency can cause pale green or yellow patches in younger leaves. This condition is known as gray speck, and is characterized by necrotic spots that form in older leaves (Figure 1).

Figure 1: Soybean leaves with an adequate (A) and deficient (B-C-D) supply of Mn (Pictures: I. Cakmak)

Manganese deficiency symptoms in corn and wheat (Pictures: IPNI).
Physiological Functions
Manganese plays a key role in photosynthesis, as the photosystem II-water oxidizing system has an absolute Mn requirement (Hakala et al. 2006). Adequate Mn is critical in this system, as Mn facilitates the photolysis (light splitting) of water molecules and provides energy for photosynthesis. It is, therefore, not surprising that Mn deficiency substantially impairs photosynthesis, even in the absence of visual leaf symptoms. The negative effect of Mn deficiency in photosynthesis results in marked decreases in soluble sugar concentrations in different parts of plants (Figure 2). It is widely believed that the reduction in photosynthesis is the major reason behind the decline in dry matter production and yield under Mn-deficient conditions.
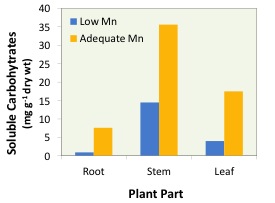
Figure 2: The influence of low and adequate Mn supply on soluble carbohydrate concentrations observed in the roots, stems and leaves of bean plants (redrawn from Marschner, 2012).
Tolerance to Pathogenic Infection
Lignin Synthesis
As a cofactor, Mn is reported to activate over 35 enzymes, several of which catalyze different steps of the lignin and phytoalexins biosynthesis. Impairment of lignin biosynthesis in Mn-deficient plants, especially in the roots, is associated with increased pathogenic attack, particularly soil-born fungi, because lignin serves as a barrier against pathogenic infection (Figure 3; Marschner 2012). Manganese application contributes to the resistance against not only various soil-borne diseases including take-all in wheat, common scab in potato and root rot in cotton, but also fungal leaf diseases such as tan spot in wheat, powdery mildew in grape and black leaf mold in tomato (Brennan 1992; Graham and Webb, 1991; Heine et al. 2011; Yao et al. 2012).
Peroxidase Enzyme
The peroxidase enzyme, which generates hydrogen peroxide, is another Mn-dependent enzyme that contributes to pathogen resistance. The hydrogen peroxide produced is not only involved in the stabilization of the cell wall, but is also thought to be directly toxic to pathogens (Heine et al. 2011), and therefore acts as a fungicide (Graham and Webb, 1991).
Stress Tolerance
Nearly all environmental stress factors represent an oxidative stress. Manganese plays an important role in improving stress tolerance, as superoxide dismutase enzymes, which are responsible for the detoxification of the destructive free radicals, require different metal cofactors, such as Mn, to function. Not surprisingly, increases in activity of Mn-superoxide dismutase contributed greatly to plant tolerance of different environmental stress factors such as winter hardiness, ozone stress, salinity and drought stress.
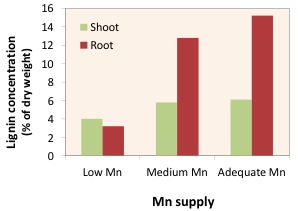
Figure 3: Effect of lignin concentration in the shoots and roots of wheat plants, as a function of Mn supply (redrawn from Marschner, 2012).
Manganese deficiency in plant tissues has also been reported to impair fatty acid production, which can adversely affect the cuticular wax deposition, as wax synthesis begins with fatty acid synthesis in plastids. Since the wax layer is responsible for limiting non-stomatal water loss and reducing the heat load on leaves (Hebbern et al. 2009), weakening of this layer due to Mn deficiency can cause an increase in the susceptibility of crops to both drought and heat stress. In barley, for example, latent Mn deficiency was found to significantly reduce the wax content (up to 40%), resulting in increased transpirational water loss and lower water-use-efficiency (Hebbern et al. 2009).
Conclusion
To sum up, prevention and correction of Mn deficiency by fertilization can:
- Enhance the photosynthetic efficiency and dry matter production
- Provide resistance to biotic stress by increasing plant resistance to various diseases and reducing the need for fungicides
- Contribute to abiotic stress tolerance, particularly drought and heat
- Result in significantly higher crop yield.
Readings
Alloway BJ. (2008) Micronutrients and Crop Production: An Introduction. In: Alloway BJ (Ed) Micronutrient Deficiencies in Global Crop Production. Springer, pp 1-40
Brennan RF (1992) The role of manganese and nitrogen nutrition in the susceptibility of wheat plants to take-all in western Australia. Fertilizer Res 31:35-41
Graham RD, Webb MJ (1991) Micronutrients and plant disease resistance and tolerance in plants. In Micronutrients in Agriculture (J. J. Mortvedt, F. R. Cox, L. M. Shuman and R. M. Welch, eds.), pp. 329–370. SSSA Book Series No. 4, Madison, WI.
Hakala M, Rantamaki S, Puputti EM, Tyystjarvi T, Tyystjarvi E (2006) Photoinhibition of manganese enzymes: insights into the mechanism of photosystem II photoinhibition. J Exp Bot 57:1809-1816
Hebbern CA, Laursen KH, Ladegaars AH, Schmidt SB, Pedas P, Bruhn D, Schjoerring JK, Wulfsohn D, Husted S (2009) Latent manganese deficiency increases transpiration in barley ( Hordeum vulgare). Physiol Plant 135:307-316
Heine, G., J.F.J. Max, H. Fuhrs, D. W. Moran-Puente, D. Heintz, and W. J. Horst. (2011) Effect of manganese on the resistance of tomato to Pseudocercospora fuligena. J. Plant Nutr. Soil Sci. 174:827-836.
Marschner P (2012) Marschner’s Mineral Nutrition of Higher Plants (3rd ed). Elsevier Science
Yu Q, Osborne L, Rengel Z (1998) Micronutrient deficiency changes activities of superoxide dismutase and ascorbate peroxidase in tobacco plants. J Plant Nutr 21:1427-1437